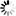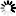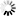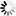Seebio(R) 烟酰胺单核苷酸应用
Seebio®烟酰胺单核苷酸(nicotinamide mononucleotide,NMN)是一种自然存在的生物活性核苷酸,NMN有2种不规则存在形式,α和β;β异构体是NMN的活性形式,分子量为334.2g/mol。
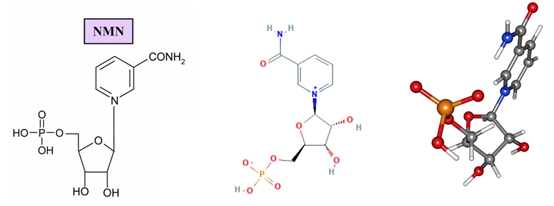
烟酰胺单核苷酸化学结构式和球棍模型
在细胞内,尼克酰胺单核苷酸(NMN)是烟酰胺腺嘌呤二核苷酸(NAD+)的重要前体。NAD+是一种在细胞内参与多种生物化学反应的辅酶,对于维持细胞正常功能和代谢至关重要。因此,NMN在细胞内具有重要的生物学意义。
|
产品名称
|
CAS
|
纯度
|
包装
|
|
β-烟酰胺单核苷酸 (NMN)
|
1094-61-7
|
99%
|
1kg 5kg 10kg 25kg
|
详询西宝生物产品,请咨询:400-021-8158 / 021-50272975。
NMN对NAD+的促进作用
主要体现在其作为NAD+的前体的功能上。在合成途径中,烟酰胺核糖或烟酰胺通过NRK(烟酰胺核苷激酶)或NAMPT、NMNAT合成烟酰胺单核苷酸(NMN),随后,NMN通过NMNAT1-3酶合成NAD+。
尽管不能在血清中检测到完整结构的NMN,但摄入NMN仍能够在很短的时间内(15分钟)显著提高雌性和雄性小鼠的NAD+水平。
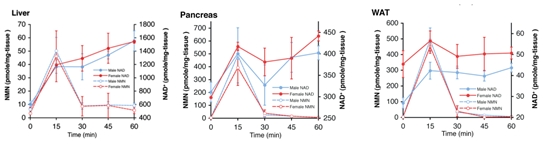
肝脏、胰腺、白色脂肪组织NMN、NAD+水平
NMN功效
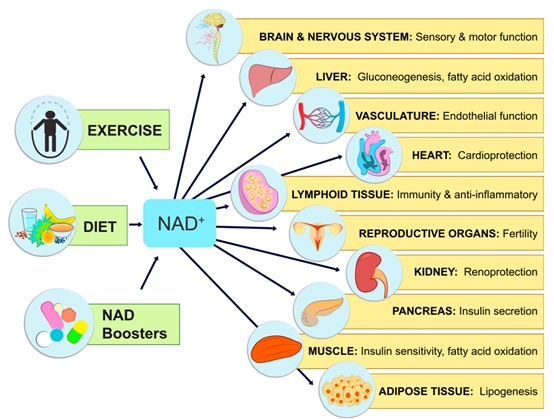
NMN的主要作用是通过转化为NAD+发挥其生物学功能。NAD+,全名烟酰胺腺嘌呤双核苷酸,又被称为辅酶I,广泛分布在人体的所有细胞内,参与上千种生物催化反应,是人体内不可或缺的辅酶。
在衰老过程中,NAD+水平的下降被认为是导致各种疾病和残疾的主要原因,包括抗衰老、认知和运动功能障碍、免疫缺陷,以及由自身免疫炎症反应失调引起的关节炎、代谢障碍和心血管疾病。
通过补充NMN来提高体内NAD+含量,有助于延缓、改善和预防与衰老相关的多种表型,包括年龄诱导的代谢紊乱和老年疾病。
NAD+与衰老
人体持续进行能量合成与代谢,以保持生命活动的平衡与健康。在这一过程中,烟酰胺腺嘌呤二核苷酸(NAD+)作为氧化还原反应的辅酶,扮演着关键角色。NAD+直接或间接地调控多个细胞功能,包括线粒体内的能量合成、DNA损伤修复、染色质重塑,以及细胞衰老和免疫细胞功能。这些细胞的基本生理过程和功能对于维持组织、代谢动态平衡及健康衰老至关重要。
衰老是人体的自然过程。随着年龄增长,人体内NAD+水平逐渐减少,导致多个器官内的线粒体能量合成下降,包括大脑、脂肪组织、皮肤、肝脏、骨骼肌和胰腺。与NAD+水平的降低相关联的是一系列与年龄有关的疾病,如认知功能减退、癌症、代谢性疾病、肌少症和衰弱。因此,通过调节NAD+代谢通路,增加NAD+水平可能成为改善衰老相关疾病、延长人类寿命和健康寿命的潜在途径。
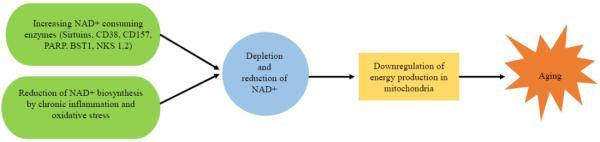
衰老过程中体内NAD+减少的原因
NMN与抗衰老
作为NAD+的前体物质,NMN理论上通过增加体内NAD+水平,减缓随年龄增长而导致的NAD+枯竭过程,从而成为一种抗衰老保健品。
科学界通过细胞试验、动物模型和临床试验对NMN的抗衰老活性进行了广泛研究。研究发现,NMN在啮齿动物模型和人体水平上均表现出有益效果,如保护内皮细胞、改善血供、改善代谢功能障碍以及保护神经系统。一项2021年的随机双盲临床试验显示,NMN治疗对绝经后超重或肥胖的糖尿病前期女性产生了积极效果,增强了肌肉对胰岛素的敏感性。类似的研究在2023年也表明,NMN治疗能够显著提高血清烟酰胺水平,减缓动脉僵硬度。此外,NMN的长期使用和潜在的副作用需要更多深入的研究和验证。
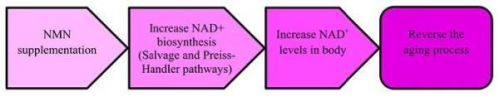
NMN抗衰老的机制
NAD+与神经系统
Sirtuins是依赖NAD+的脱酰基酶,传统上与热量限制和衰老有关。在神经发育中,SIRT1通过Akt-GSK3通路促进轴索生长、神经突生长和树枝状分支。在突触的发育和调节中,sirtuins蛋白在生理和损伤后都发挥着重要的调节作用。特别是在海马体中,SIRT1以抑制型复合体形式存在,参与调控microRNA-134的转录因子YY1,这对于突触的形成和长期的增强至关重要。
在神经疾病的发展中,SIRT1发挥着保护作用,尤其在阿尔茨海默氏病、帕金森氏病和运动神经元病等神经退行性疾病中。这些保护作用可能与SIRT1在代谢、抗应激和基因组稳定性方面的功能有关。因此,激活SIRT1的药物可能为治疗这些神经疾病提供一种有希望的方法。
NAD+与癌症
研究表明增加NAD+水平可能对癌症治疗具有潜在效果。NMNAT3的过表达可以提高线粒体NAD+水平,从而抑制胶质母细胞瘤细胞的生长。补充烟酰胺(NA)或烟酰胺单核苷酸(NAM)可以抑制SCID小鼠的肿瘤生长和多器官肿瘤转移。其原理包括过量的NAD+促进线粒体呼吸,降低糖酵解,抵消癌细胞倾向的Warburg代谢。增加NAD+还能活化SIRT1和SIRT6,两者通过下调β-catenin信号和抑制糖酵解来抑制肿瘤。然而存在一些矛盾和担忧,如NAD+促进DNA修复和血管生成,可能有助于癌细胞的生长。此外,降低肿瘤NAD+水平可能增加癌细胞对化疗药物的敏感性。因此,在标准癌症模型中进一步测试NAD+补充剂的效果是非常重要的。
NAD+与昼夜节律
NAD+是一种关键的生物分子,其依赖的脱乙酰酶SIRT1通过连接调节NAD+补救途径的酶反馈回路和昼夜节律转录-翻译反馈回路,成为昼夜节律与代谢之间的桥梁。NAD+通过SIRT1实现对生物钟的调节,其中SIRT1将BMAL1和PER2去乙酰化,抑制CLOCK-BMAL1介导的clock genes的转录。此外,NAD+通过影响SIRT1的去乙酰化活性,反过来影响包括NAMPT在内的一系列生物钟相关蛋白的表达。
生物钟调节与多种疾病相关,包括但不限于睡眠障碍、糖尿病和肿瘤。紊乱的生物钟可能是多种病理过程的触发因素,可能源自遗传或环境因素。总体而言,维持正常的生物钟功能对于保持健康至关重要。
NAD+与肝功能
NAD+信号通路中的酶已被证明可以保护肝脏免受脂肪堆积、纤维化和胰岛素抵抗的影响,这些因素与脂肪肝病发生有关。NAMPT在高脂饮食诱导的脂肪肝发生中发挥关键调节作用,其抑制将使肝脂肪变性更加严重,而NAMPT的过表达则改善肝脂质积累。SIRT1及其下游靶点(PGC-1a、PSK9和SREBP1)维持线粒体功能、胆固醇转运和脂肪酸稳态。SIRT2通过去乙酰化磷酸烯醇丙酮酸羧激酶来控制糖异生,SIRT3调控OXPHOS、脂肪酸氧化、酮生成和抗氧化应激,而SIRT6控制糖异生。由于这些通路在肝脏中的重要性,维持NAD+水平对于维持器官功能至关重要。在正常情况下,由于肥胖和衰老,NAMPT水平下降,CD38水平升高,导致到中年时,稳态NAD+水平下降2倍。将NAD+水平提高到年轻水平在预防和治疗肥胖、酒精性脂肪性肝炎和NASH方面表现出显著效果,同时还改善葡萄糖稳态和线粒体功能,增强肝脏再生能力,保护肝脏免受肝毒性损害。
NAD+与肾功能
老年肾脏中NAD+水平的降低和sirtuin活性的下降在很大程度上是肾功能和顺应性随年龄下降的原因。通过NAD+补充可以激活SIRT1和SIRT3,从而保护高糖诱导的肾系膜细胞肥大。同时,使用NMN治疗小鼠以SIRT1依赖的方式保护顺铂诱导的急性肾损伤(AKI)。5-氨基咪唑-4-羧胺核苷可刺激AMPK活性,增加NAD+水平,并以SIRT3依赖的方式保护顺铂诱导的AKI。小鼠补充NAM可以刺激肾脏保护前列腺素PGE2的分泌,提升缺血后肾功能;同时,NAM也通过刺激NAD+合成抑制顺铂诱导的AKI。
NAD+与骨骼肌
与年轻的野生型小鼠相比,老年小鼠的肌肉表现出萎缩、炎症标志物增加以及胰岛素信号和胰岛素刺激葡萄糖摄取能力下降。有关骨骼肌的研究表明:用NAD+前体治疗老年小鼠可以显著改善肌肉功能。使用NMN进行治疗可以通过增加线粒体功能、提高ATP生成、减少炎症,并将糖酵解II型肌肉转变为氧化纤维型肌肉,逆转与年龄相关的有害变化。这表明NAD+的补充可能有助于保持肌肉健康和功能,特别是在老年阶段。
NAD+与心脏功能
NAD+水平对正常心脏功能和损伤后的恢复至关重要。有关心脏功能的研究表明:SIRT3在心脏功能中是至关重要的,SIRT3敲除小鼠表现为OXPHOS酶的高度乙酰化,导致ATP减少,使主动脉对收缩高度敏感,可能与线粒体通透性过渡孔的调节因子CypD的激活有关。SIRT3-KO鼠在13个月大时出现纤维化和心肌肥厚,随着年龄增长,病情进一步加剧,而NMN治疗可以逆转这种下降。使用NAMPT过表达或NMN治疗能显著防止压力过载和缺血-再灌注损伤,减少梗死面积。NAD+前体治疗改善了老年MDX心肌病小鼠的心脏功能。NAD+前体提高了缺铁诱导的心力衰竭小鼠模型的线粒体和心脏功能。NAD+前体通过激活SIRT3保护并恢复弗里德希氏共济失调(FRDA)心肌病小鼠模型的心脏功能。
NAD+与血管内皮细胞
内皮细胞(EC)的衰老与血管疾病密切相关,而NAD+前体NMN的补充在一些研究中显示了改善效果:NMN治疗老年小鼠可恢复颈动脉内皮依赖性扩张,降低主动脉脉搏波速和弹性动脉刚度。NMN对老年小鼠的治疗取得显著疗效,通过促进SIRT1-依赖的毛细血管密度的增加,改善了老年小鼠的血液流动和耐力。NMN通过改善老年小鼠的血管内皮功能和神经血管耦合反应,显著提高老年小鼠的认知。同时,NMN还降低了老龄鼠脑微血管内皮细胞的线粒体ROS,恢复了NAD+和线粒体能量。在血管内皮中增加NAD+水平可能成为一种提高老年人活动能力的潜在疗法,并能治疗因血流减少而导致的疾病,如缺血-再灌注损伤、伤口愈合缓慢、肝功能障碍和肌肉肌病等。
NAD+与代谢障碍
NMN对脂肪代谢和糖代谢紊乱导致的肥胖、Ⅱ型糖尿病、生殖抑制都具有改善作用。甚至能够改善肥胖母亲对雌性后代生殖的不良影响。 NAD+前体的补充可能在代谢障碍的治疗中发挥重要作用。
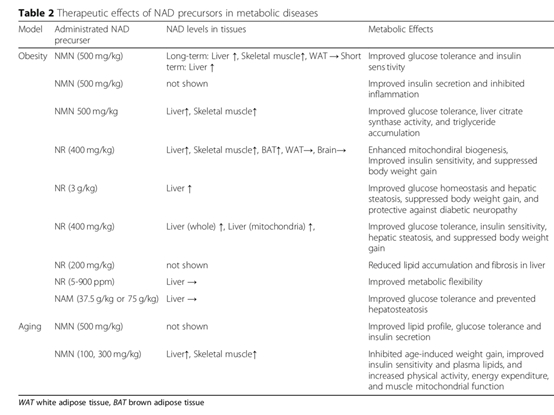
NAD+前体在动物实验中对代谢障碍的治疗作用
参考文献:
[1].Nadeeshani, H., et al., Nicotinamide mononucleotide (NMN) as an anti-aging health product - Promises and safety concerns. J Adv Res, 2022. 37: 267-278.
[2]. Liu, L., Su, X., Quinn, W. J., 3rd, Hui, S., Krukenberg, K., Frederick, D. W., . . . Rabinowitz, J. D. (2018). Quantitative Analysis of NAD Synthesis-Breakdown Fluxes. Cell Metab, 27(5), 1067-1080 e1065. doi:10.1016/j.cmet.2018.03.018
[3]. Bonkowski, M. S., & Sinclair, D. A. (2016). Slowing ageing by design: the rise of NAD(+) and sirtuin-activating compounds. Nat Rev Mol Cell Biol, 17(11), 679-690. doi:10.1038/nrm.2016.93
[4]. Clement, J., Wong, M., Poljak, A., Sachdev, P., & Braidy, N. (2019). The Plasma NAD(+) Metabolome Is Dysregulated in "Normal" Aging. Rejuvenation Res, 22(2), 121-130. doi:10.1089/rej.2018.2077
[5]. Yang, Y., & Sauve, A. A. (2016). NAD(+) metabolism: Bioenergetics, signaling and manipulation for therapy. Biochim Biophys Acta, 1864(12), 1787-1800. doi:10.1016/j.bbapap.2016.06.014
[6].H. Kim, E.L. Jacobson, M.K. Jacobson, Science 261 (1993) 1330–1333.
[7]. Pehar, M., Harlan, B. A., Killoy, K. M., & Vargas, M. R. (2018). Nicotinamide Adenine Dinucleotide Metabolism and Neurodegeneration. Antioxid Redox Signal, 28(18), 1652-1668. doi:10.1089/ars.2017.7145
[8].M.B. Schultz, D.A. Sinclair, Cell Metab. 23 (2016) 965–966.
[9]. Davila, A., Liu, L., Chellappa, K., Redpath, P., Nakamaru-Ogiso, E., Paolella, L. M., . . . Baur, J. A. (2018). Nicotinamide adenine dinucleotide is transported into mammalian mitochondria. Elife, 7. doi:10.7554/eLife.33246
[10].M. Howard, J.C. Grimaldi, J.F. Bazan, F.E. Lund, L. Santos-Argumedo, R.M. Parkhouse, T.F. Walseth, H.C. Lee, Science 262 (1993) 1056–1059.[6
[11].J. Camacho-Pereira, M.G. Tarragó, C.C.S. Chini, V. Nin, C. Escande, G.M. Warner, A.S. Puranik, R.A. Schoon, J.M. Reid, A. Galina, E.N. Chini, Cell Metabolism 23 (2016) 1127–1139.
[12]. Chini, C. C. S., Tarrago, M. G., & Chini, E. N. (2017). NAD and the aging process: Role in life, death and everything in between. Mol Cell Endocrinol, 455, 62-74. doi:10.1016/j.mce.2016.11.003
[13]. Grozio, A., Mills, K. F., Yoshino, J., Bruzzone, S., Sociali, G., Tokizane, K., . . . Imai, S. I. (2019). Slc12a8 is a nicotinamide mononucleotide transporter. Nat Metab, 1(1), 47-57. doi:10.1038/s42255-018-0009-4
[14]. Araki, T., Sasaki, Y., & Milbrandt, J. (2004). Increased nuclear NAD biosynthesis and SIRT1 activation prevent axonal degeneration. Science, 305(5686), 1010-1013. doi:10.1126/science.1098014
[15]. Ratajczak, J., Joffraud, M., Trammell, S. A., Ras, R., Canela, N., Boutant, M., . . . Canto, C. (2016). NRK1 controls nicotinamide mononucleotide and nicotinamide riboside metabolism in mammalian cells. Nat Commun, 7, 13103. doi:10.1038/ncomms13103
[16]. Pehar, M., Harlan, B. A., Killoy, K. M., & Vargas, M. R. (2018). Nicotinamide Adenine Dinucleotide Metabolism and Neurodegeneration. Antioxid Redox Signal, 28(18), 1652-1668. doi:10.1089/ars.2017.7145
[17].J. Clement, M. Wong, A. Poljak, P. Sachdev, N. Braidy, Rejuvenation Res. (2018).
 |
 |
 |
| 官网:www.cxbio.com | 微信服务号:iseebio | 微博:seebiobiotech |
 |
 |
 |
| 商城:mall.seebio.cn | 微信订阅号:seebiotech | 泉养堂:www.canmedo.com |
相关资讯
- Cell:体内唾液酸化作用可将有害抗体变为有益抗炎抗体
- Cell:鉴定出弥漫大B细胞淋巴瘤的驱动基因
- 天野脂肪酶Amano Lipase-符合“绿色化学”观念的生物催化剂
- 乳糖/D-半乳糖[快速]检测试剂盒
- 西宝生物 Jackson Immuno Research - 代理证书
- 科学家发现抗高温基因 水稻“高温不死”还能增产
- 增塑剂让快餐不健康 我们还是去“吃土”吧!
- Nature:重磅!一些人胚胎干细胞系发生癌症相关突变
- PNAS:解读!科学家们有望利用一种新型的聚糖标记策略来揭秘肝脏中的免疫激活机制!
- Culture(R) A-83-01


新进产品
同类文章排行
- 羟丙基壳聚糖:性能提升与多领域应用
- “杂草”变身宝贝——油莎草根油 沙漠中的软黄金
- 青刺果油:源自自然的保湿抗衰产品
- 虾青素功效及应用场景
- 玻色因原料: 全方位修复皮肤的抗衰剂
- 谷胱甘肽功效及应用场景
- 辅酶Q10功效及应用场景
- 花生四烯酸(Arachidonic acid)功能及应用场景
- 黑曲霉糖(Nigerose)功效及应用场景
- 曲二糖 (Kojibiose,CAS: 2140-29-6)性质及应用场景
资讯文章
您的浏览历史